被子植物是植物界中最大和最高级的一类,全世界约有300~450个科属。由少数(典型的有8个)细胞构成的胚囊和双受精现象被视为被子植物在进化上的一致性和与其他植物类群区别的证据。本文主要为你介绍的是被子植物的额生殖结构和发育过程。

被子植物的生殖结构与发育
一、被子植物的雄性生殖器官的结构与功能:
1.花药的结构;
药室内壁通常仅一层细胞,初期常贮藏大量物质。花药成熟时,细胞径向扩展,贮藏物质消失,除外切向壁外,均发生多条斜纵向条纹状的纤维素,次生加厚,故称纤维层。药室内壁在形成纤维层时,常在两个相邻花粉囊交接处留下一个狭长的薄壁细胞――裂口。
中层由一至数层较小的细胞组成,初期可贮存有淀粉等营养物质,以后其细胞被挤压逐渐解体和被吸收,所以,其在成熟的花药中已消失。
绒毡层的细胞较大, 初期为单核,但在花粉母细胞开始减数分裂时,形成双核或多核。该层胞质浓,胞器丰富,胞质含较多的RNA及蛋白质,并含有丰富的油脂和类胡萝卜素等营养物质,对花粉粒的发育或形成起着重要的营养和调节作用,后渐退化。绒毡层能合成和分泌胼胝质酶,分解花粉母细胞和四分体的胼胝质壁,使单核花粉粒分离。绒毡层还合成蛋白质,通过运转至花粉粒的外壁上,这是一种识别蛋白。
花粉粒成熟后,纤维层细胞失水,所产生的机械力使花药在裂口处断开,花粉粒由裂口处纵轴形成的裂缝散出。花粉囊壁因绒毡层的解体而消失,或仅存痕迹,只剩有表皮及纤维层。
2.花药的发育过程;
花药由花粉囊和药隔组成。花药发育初期,结构简单,外层为一层原表皮,内层为一群基本分生组织;不久,由于花药四个角隅处分裂较快,花药呈四棱形。以后在四棱处的原表皮下面分化出多列体积较大,核亦大,胞质浓,径向壁较长,分裂能力较强的孢原细胞。随后孢原细胞进行平周分裂,成内、外两层,外层为初生周缘层;内层为初生造孢细胞,初生周缘层细胞继续平周分裂和垂周分裂,逐渐形成药室内壁、中层及绒毡层。花药中部的细胞逐渐分裂,分化形成维管束和薄壁细胞,构成药隔。
3.小孢子的产生;
造孢细胞经几次分裂后,形成花粉母细胞。花粉母细胞排列紧密,早期具有一般的纤维素壁。整个花粉囊内,花粉母细胞之间与绒毡层细胞之间都有胞间连丝相贯通。在相邻的花粉母细胞间,常形成直径为1~2微米的胞质管,将同一花粉囊的花粉母细胞连接成为合胞体。研究发现有染色质、内质网片段等细胞器、营养物质等通过胞质管进行物质交流。这种连接现象同花粉囊中花粉母细胞的减数分裂同步化,营养物质、生长物质的迅速运输及分配有关。
以后,在减数分裂过程中,随着花粉母细胞在质膜与细胞壁之间积累胼胝质,形成胼胝质壁,并逐渐加厚,致使胞间连丝和胞质管被阻断。花粉母细胞在减数分裂前,细胞的形态和结构常有很多变化,如细胞和核的体积增大,胞质增稠,RNA和蛋白质大量合成,核蛋白体密集,质体和线粒体增多,液泡小而不明显等,所以花粉母细胞显著地与周围花粉囊壁细胞不同。
4.花粉(雄配子体)的发育过程;
初生造孢细胞――花粉母细胞――四分体――单细胞花粉粒――二细胞花粉(营养细胞、生殖细胞)――三细胞花粉(营养细胞1个,精子3个)
5.成熟花粉粒的结构与功能
花粉壁、营养细胞、生殖细胞、精子。
成熟花粉粒与蜜蜂
二、雌性生殖器官的结构与发育:
1.雌蕊的结构;
雌蕊由心皮构成,心皮有3条维管束,与叶脉相当的为背束,其两侧的维管束为腹束。心皮向内卷合,使近轴的一面(或称腹面)闭合起来,心皮边缘连合之处称为腹缝线,其背束之处称为背缝线。单雌蕊只有一条腹缝线和背缝线。在复雌蕊或合生心皮雌蕊中,其腹缝线和背缝线的数目与心皮数目相同。心皮卷合成雌蕊后,其上端为柱头,中间为花柱,下端为子房。
2.成熟胚珠的结构;
合点:珠心基部,珠被、珠心和珠柄连合的部位。
珠心:胚珠中最重要的部分,位于珠被内的薄壁组织,其中产生大孢子,进一步发育成胚囊。
珠被:包被珠心组织的结构,有的植物可有外珠被和内珠被。
珠孔:珠被前端未闭合留下的小孔。
珠柄:胚珠基部着生于胎座上的小柄,维管束经胎座进入胚珠。

3.胚珠的发育;
胚珠的发育过程首先在胎座表皮下层的一些细胞经平周分裂,产生一团突起,成为胚珠原基。原基的前端成为珠心。珠心是一团相似的薄壁细胞,原基的基部成为珠柄。由于珠心基部的表皮层细胞分裂较快,产生一环状的突起,逐渐向上生长扩展,将珠心包围,此包围珠心的组织即为珠被,仅在珠心的前端留下一小孔,即珠孔。
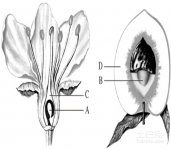
4.胚囊的结构与发育;
胚囊是被子植物的雌配子体,由卵细胞、助细胞、极核和反足细胞组成的结构。
卵细胞是一个有高度极性的细胞,通常它的壁在珠孔端最厚,接近合点端的壁逐渐变薄。卵细胞核大,核仁的RNA含量高于胚囊中其他细胞,细胞内常有一个大液泡。核和细胞质等在卵细胞内的分布也常有明显的极性;成熟卵细胞中,质体和线粒体常退化,数量减少,内质网及高尔基体也常变得稀少或不发达,反映出卵细胞代谢活动的强度是低的。
助细胞与卵细胞在珠孔端排列成三角形。它们也是有高度极性的细胞。助细胞的壁和卵细胞一样,也以珠孔端为最厚,向合点端逐渐变薄。助细胞最突出的特征是在珠孔端的细胞壁上有丝状器结构,它是壁向内延伸的部分,类似传递细胞壁的内褶突起。不同植物的组成常有变化。丝状器的结构,大大地增加了质膜的表面面积,这可能与助细胞的功能有关。
中央细胞是胚囊中最大的一个细胞,也是高度液泡化的细胞。成熟胚囊的增大,主要由于中央细胞液泡的膨大。中央细胞的壁厚薄变化很大,在与卵细胞和中央细胞相接处,通常只有质膜而没有细胞壁;而与反足细胞相接处,则有胞间连丝的薄壁。不少植物中央细胞壁的内侧也有许多指状的内突,说明它能从珠心组织或珠被组织吸取营养物质,中央细胞的细胞质内在不少植物中发现含丰富的质体、核蛋白体、线粒体、高尔基体和内质网。中央细胞的极核或次生核常被许多细胞索悬挂在高度液泡化的细胞中间,极核通常很大,核仁也很大。
反足细胞是胚囊中一群变异最大的细胞,不仅细胞数目差异很大,在细胞的结构上也因植物而有各种变化。反足细胞具有从珠心吸收营养物质,经过反足细胞输入中央细胞的功能。大多数植物反足细胞的细胞质含丰富的质体、核蛋白体、线粒体、高尔基体和内质网。反足细胞是代谢活动非常活跃的细胞,对胚囊的发育具有吸收、传输和分泌营养物质的多种功能。除一些植物的反足细胞能存在较长的时间外,在多数植物中,它们通常是短命的,在受精前或受精后不久退化。
三、胚囊的发育:
首先,在胎座表皮下层的一些细胞经平周分裂,产生一团突起,成为胚珠原基。原基的前端成为珠心。珠心是一团相似的薄壁细胞。原基的基部成为珠柄。
通常在靠近珠孔一端的表皮下,逐渐形成一个与周围不同的细胞,即孢原细胞。孢原细胞的体积较大,细胞质较浓,细胞器丰富,RNA和蛋白质含量高,液泡化程度低,细胞核大而显著,壁上具有很多胞间连丝。在分化时,核内DNA合成至四倍体或4C水平。孢原细胞在形成后,需要进一步发育长大成胚囊母细胞。其发育的形式随植物不同而有差异。很多被子植物,包括棉花等作物,其孢原细胞先进行一次平周分裂,形成内、外两个细胞: 外侧的一个称为周缘细胞,内侧的一个称为造孢细胞。周缘细胞继续进行平周分裂和垂周分裂,增加珠心的细胞层数,而造孢细胞则长大形成胚囊母细胞。
胚囊母细胞进行减数分裂的第一次分裂:由于珠心基部的表皮层细胞分裂较快,产生一环状的突起,逐渐向上生长扩展,将珠心包围,仅在珠心的前端留下一孔。此包围珠心的组织即为珠被。
减数分裂的二次分裂:形成四分体。近珠孔端的3个细胞退化。近合点端的细胞逐渐发育为单核胚囊。并从珠心组织中不断吸取营养物质,体积不断增大,细胞核也稍有增大,同时出现大液泡。以后单核胚囊进行三次有丝分裂形成八核胚囊。
第一次分裂,分别移到胚囊细胞的两端,形成二核胚囊。第二次分裂,形成四核胚囊。胚囊细胞进一步长大,核暂时游离于共同的细胞质中。第三次分裂,形成八核胚囊。随后,每一端的四核中,各有1核向胚囊的中部移动,互相靠拢。
成熟的八核胚囊,一般珠被前端会留下一小孔,即珠孔。

")